El monte serrano
En cuanto a extensión, el monte serrano es la segunda formación boscosa de importancia en el país luego del monte ribereño. Se trata de una formación que se desarrolla en áreas predregosas que incluyen cerros, sierras y asperezas, normalmente asociados a cursos de agua y manantiales.Si bien es un tipo de monte muy modificado por el ser humano, normalmente se caracteriza por una vegetación relativamente alta en las faldas, que va disminuyendo en altura a medida que asciende, siendo sus árboles sustituidos por arbustos al aproximarse a la cima, normalmente ocupada por vegetación herbácea. Si bien en la actualidad es un monte relativamente bajo, es necesario remarcar que esa no es una característica típica del monte, sino el resultado de las actividades de corta llevadas a cabo desde hace muchos años. Por ejemplo, en los actuales montes achaparrados de la zona de Pan de Azúcar antiguamente se podía transitar bajo los mismos a caballo, según aseguran viejos pobladores de la zona. Ello se debe a que los árboles nacidos de semilla normalmente tienen un solo fuste, en tanto que los rebrotes de árboles cortados están constituidos por varios fustes más bajos, lo que convierte a un monte alto en otro achaparrado.
Este tipo de monte cumple una función primordial en la conservación de las cuencas hídricas, dado que se ubica en las nacientes de prácticamente todos los cursos de agua que tienen su origen en nuestro territorio. Siendo que a su vez se desarrolla sobre suelos con pendientes pronunciadas, su presencia es vital para evitar la erosión. Pese a ello, la superficie ocupada por el monte serrano ha disminuido sensiblemente, habiendo siendo sustituido o por praderas (salpicadas de los pocos árboles que lograron sobrevivir) o por plantaciones de eucaliptos que afectan negativamente el funcionamiento hidrológico de las cuencas.
Pero el monte serrano no sólo ha disminuido sustancialmente en superficie, sino que además ha sido profundamente modificado en cuanto a su composición de especies vegetales. Ambos procesos (disminución en superficie y en especies) han generado a su vez graves impactos sobre las especies de fauna que de él dependen.

El monte ribereño
Este tipo de bosque se ubica acompañando los principales espejos de
agua del país y se desarrolla tanto en las márgenes de los cursos de
agua, como en las de las lagunas e islas vinculadas a los mismos.
En el caso específico de los cursos de agua y lagunas del este del litoral sur, el bosque desaparece a medida que aumenta la salinidad, por lo que en su desembocadura en el océano se encuentran desprovistos de árboles.
El ancho del monte a ambos lados de los cursos de agua parece guardar relación directa con el relieve del terreno, que determina a su vez el área de inundación de la cuenca.
En general, los montes que se ubican en llanuras de inundación amplias (como el caso del río Cebollatí) tienen un ancho considerable, que puede llegar a varios cientos de metros, en tanto que aquellos ubicados en ríos más encajonados tienden a ser de un ancho de cien o menos metros. Igualmente, el ancho de los montes en general es mayor en los sitios en que un curso de agua desemboca en otro y el caso más notorio es el del llamado Rincón de los Gauchos, donde se unen los ríos Queguay Grande y Queguay Chico, dando lugar a uno de los montes más extensos del país.
A su vez, existen importantes diferencias entre los montes ribereños del noreste y noroeste con los del centro y sur del país, donde los primeros se caracterizan por la presencia de especies de gran talla y de una fauna más diversa y abundante. Ello se debe a que muchas de esas especies vegetales y animales parecen alcanzar allí el límite sur de su dispersión, a lo que se suma una menor presencia humana que en los más poblados centro y sur.
La vegetación arbórea y arbustiva de este tipo de monte se desarrolla en general siguiendo un patrón similar, determinado en primer lugar por las necesidades hídricas de cada especie, a lo que se suman además sus necesidades edáficas (de suelo) y lumínicas.
En términos generales, el monte se dispone en tres franjas paralelas al curso de agua. Contra el margen se establecen aquellas con mayores requerimientos hídricos, tales como sarandíes, sauces, mataojos. Estas especies juegan un papel esencial en la conservación del curso de agua. Por un lado, protegen a las márgenes de la erosión, fijándolas con sus raíces y protegiéndolas en las inundaciones con su ramaje. Por otro lado, algunas de ellas (en particular los sarandíes) contribuyen a disminuir la velocidad del agua, con lo cual también ayudan a la conservación de la cuenca.
En la franja intermedia del monte se ubican aquellas especies con requerimientos hídricos menores a las anteriores, pero que a su vez no se adaptan a condiciones de sequía o a condiciones extremas de temperatura. Si bien muchas de ellas requieren bastante luminosidad, la mayoría se adapta al ambiente sombrío del monte durante su etapa juvenil y sólo llega al estrato superior en su estado adulto. La composición de especies varía bastante de un lugar a otro, pero es aquí donde en general se encuentran las especies de mayor porte como los laureles, azoita cavalo, tarumán, tembetarí, etc.
Finalmente, contra el borde exterior del monte se instalan las especies mejor adaptadas a resistir los extremos de temperatura, los vientos y la escasez de agua, que en general requieren a su vez de un nivel elevado de luminosidad. Entre las especies típicas se encuentran los molles, arueras, canelones.
Pese a lo antedicho, en muchos montes del país suele ocurrir que tanto contra el margen del curso de agua como en la zona externa del monte se encuentren las especies típicas de la zona intermedia. Ello no es un hecho fortuito, sino el resultado de la acción humana, que ha provocado cambios profundos en el régimen de muchos cursos de agua del país. Como resultado, las barrancas han sido erosionadas por las aguas, determinando la desaparición de las especies que allí se ubicaban, por lo que las de la zona intermedia han quedado contra el margen. En la zona externa, tanto las actividades de tala como el afán
de agrandar los campos han significado la desaparición de la franja externa, por lo que también aquí las especies de la zona intermedia han quedado como límite externo del monte.
PRADERAS.
En el caso específico de los cursos de agua y lagunas del este del litoral sur, el bosque desaparece a medida que aumenta la salinidad, por lo que en su desembocadura en el océano se encuentran desprovistos de árboles.
El ancho del monte a ambos lados de los cursos de agua parece guardar relación directa con el relieve del terreno, que determina a su vez el área de inundación de la cuenca.
En general, los montes que se ubican en llanuras de inundación amplias (como el caso del río Cebollatí) tienen un ancho considerable, que puede llegar a varios cientos de metros, en tanto que aquellos ubicados en ríos más encajonados tienden a ser de un ancho de cien o menos metros. Igualmente, el ancho de los montes en general es mayor en los sitios en que un curso de agua desemboca en otro y el caso más notorio es el del llamado Rincón de los Gauchos, donde se unen los ríos Queguay Grande y Queguay Chico, dando lugar a uno de los montes más extensos del país.
A su vez, existen importantes diferencias entre los montes ribereños del noreste y noroeste con los del centro y sur del país, donde los primeros se caracterizan por la presencia de especies de gran talla y de una fauna más diversa y abundante. Ello se debe a que muchas de esas especies vegetales y animales parecen alcanzar allí el límite sur de su dispersión, a lo que se suma una menor presencia humana que en los más poblados centro y sur.
La vegetación arbórea y arbustiva de este tipo de monte se desarrolla en general siguiendo un patrón similar, determinado en primer lugar por las necesidades hídricas de cada especie, a lo que se suman además sus necesidades edáficas (de suelo) y lumínicas.
En términos generales, el monte se dispone en tres franjas paralelas al curso de agua. Contra el margen se establecen aquellas con mayores requerimientos hídricos, tales como sarandíes, sauces, mataojos. Estas especies juegan un papel esencial en la conservación del curso de agua. Por un lado, protegen a las márgenes de la erosión, fijándolas con sus raíces y protegiéndolas en las inundaciones con su ramaje. Por otro lado, algunas de ellas (en particular los sarandíes) contribuyen a disminuir la velocidad del agua, con lo cual también ayudan a la conservación de la cuenca.
En la franja intermedia del monte se ubican aquellas especies con requerimientos hídricos menores a las anteriores, pero que a su vez no se adaptan a condiciones de sequía o a condiciones extremas de temperatura. Si bien muchas de ellas requieren bastante luminosidad, la mayoría se adapta al ambiente sombrío del monte durante su etapa juvenil y sólo llega al estrato superior en su estado adulto. La composición de especies varía bastante de un lugar a otro, pero es aquí donde en general se encuentran las especies de mayor porte como los laureles, azoita cavalo, tarumán, tembetarí, etc.
Finalmente, contra el borde exterior del monte se instalan las especies mejor adaptadas a resistir los extremos de temperatura, los vientos y la escasez de agua, que en general requieren a su vez de un nivel elevado de luminosidad. Entre las especies típicas se encuentran los molles, arueras, canelones.
Pese a lo antedicho, en muchos montes del país suele ocurrir que tanto contra el margen del curso de agua como en la zona externa del monte se encuentren las especies típicas de la zona intermedia. Ello no es un hecho fortuito, sino el resultado de la acción humana, que ha provocado cambios profundos en el régimen de muchos cursos de agua del país. Como resultado, las barrancas han sido erosionadas por las aguas, determinando la desaparición de las especies que allí se ubicaban, por lo que las de la zona intermedia han quedado contra el margen. En la zona externa, tanto las actividades de tala como el afán
de agrandar los campos han significado la desaparición de la franja externa, por lo que también aquí las especies de la zona intermedia han quedado como límite externo del monte.
PRADERAS.
Formación herbácea densa con poca presencia de árboles.
Es el paisaje típicamente americanos, son naturales y
de
gran extensión.
Se ubica en zonas de clima templado oceánico, templado
de
transición y frío oceánico.
Los campos, a la vista,
puedan parecer monótonos.
El factor limitante de la pradera es el agua.
El promedio anual de precipitaciones llega
a 600 mm3.
En las zonas más húmedas, donde las lluvias se
equilibran
con la evaporación desde el suelo desnudo y la transpiración de las
plantas,
abundan las hierbas altas.
A medida que las precipitaciones disminuyen, las
hierbas son
más bajas y la pradera termina convirtiéndose en estepas y
semidesiertos.
Las praeras del Río de la Plata constituyen una de las
áreas
más extendidas de praderas naturales en el mundo, abarcan una región de
70
millones de hectáreas, entre el este de Argentina, Uruguay y Río Grande
do
Sul en Brasil.
Antes de la agricultura, los campos fueron
continuamente
controlados por grandes manadas de herbívoros.
Los desechos fecales de estas manadas enriquecían el
suelo y
permitían a la naturaleza sustentar una amplia diversidad de animales
silvestres.
La mayoría de los pequeños animales de las planicies
estan
adaptados a una vida subterránea, donde las condiciones a lo largo del
año
son relativamente estables.
Las praderas se encuentran en
regiones
templadas, sin temperaturas extremas, con promedios de 17º C.
Se diferencian claramente dos
estaciones: con lluvias y con
sequías (hoy no tan notoriamente por el calentamiento global).
Vegetación:
Árboles aislados
rodeados
de hierbas cortas y tiernas.
Ecosistema en el que las
gramíneas constituyen la vegetación
dominante.
Celula
célula (del latín cellula,
diminutivo de cella, "hueco") [1]
es la unidad morfológica y funcional
de todo ser vivo. De hecho, la célula es el elemento de menor tamaño
que puede considerarse vivo.[2]
De este modo, puede clasificarse
a los organismos vivos según el número de células que posean: si sólo
tienen una, se les denomina unicelulares (como
pueden ser los protozoos o las bacterias,
organismos microscópicos); si poseen más, se les llama pluricelulares.
En estos últimos el número de células es variable: de unos pocos
cientos, como en algunos nematodos, a cientos de billones
(1014), como en el caso del ser humano. Las células suelen poseer un tamaño
de 10 µm y una masa de 1 ng,
si bien existen células mucho mayores.
La teoría celular, propuesta en 1839 por Matthias Jakob Schleiden y Theodor Schwann, postula que todos los organismos están
compuestos por células, y que todas las células derivan de otras
precedentes
. De este modo, todas las funciones vitales emanan de la
maquinaria celular y de la interacción entre células adyacentes; además,
la tenencia de la información genética, base de la herencia, en su ADN
permite la transmisión de aquella de generación en generación.[3]
nacimiento de la primera célula. Si bien existen
muchas hipótesis que especulan cómo ocurrió, usualmente se describe que
el proceso se inició gracias a la transformación de moléculas
inorgánicas en orgánicas bajo
unas condiciones ambientales adecuadas;
tras esto, dichas biomoléculas
se asociaron dando lugar a entes complejos capaces de autorreplicarse.
Existen posibles evidencias fósiles
de estructuras celulares en rocas datadas en torno a 4 o 3,5 miles de
millones de años (giga-años o Ga.).[4]
[5]
[nota 1]
Se han encontrado evidencias muy fuertes de formas de vida unicelulares
fosilizadas en microestructuras en rocas de la
formación Strelley Pool,
en Australia Occidental, con una
antigüedad de 3,4 Ga. Se trataría de los fósiles
de células más antiguos encontrados hasta la fecha. Evidencias
adicionales muestran que su metabolismo
sería anaerobio y basado en el sulfuro.[6]
Existen dos grandes tipos celulares
: las procariotas (que comprenden las células de arqueas y bacterias)
y las eucariotas (divididas tradicionalmente en animales y vegetales, si bien se incluyen además hongos y protistas,
que también tienen células con propiedades características).
Historia y teoría celular
La historia de la biología celular ha estado ligada al desarrollo tecnológico que pudiera
sustentar su estudio. De este modo, el primer acercamiento a su
morfología se inicia con la popularización del microscopios
rudimentarios de lentes compuestas en el siglo
XVII, se suplementa con diversas técnicas histológicas para microscopía óptica en
los siglos XIX y XX y
alcanza un mayor nivel resolutivo mediante los estudios de microscopía
electrónica, de fluorescencia y confocal, entre
otros, ya en el siglo XX. El desarrollo de herramientas moleculares, basadas en el manejo de ácidos nucleicos y enzimas
permitieron un análisis más exhaustivo a lo largo del siglo XX.[7
Descubrimiento

Las primeras aproximaciones al estudio de la célula surgieron en el siglo
XVII;[8]
tras el desarrollo a finales del siglo
XVI de los primeros microscopios.
[9]
Estos permitieron realizar numerosas observaciones, que condujeron en
apenas doscientos años a un conocimiento morfológico relativamente aceptable. A
continuación se enumera una breve cronología de tales descubrimientos:
- 1665: Robert Hooke publicó los resultados de sus observaciones sobre tejidos vegetales
- , como el corcho, realizadas con un microscopio de 50 aumentos construido por él mismo.
- Este investigador fue el primero que, al ver en esos tejidos unidades que se repetían a modo de celdillas de un panal, las bautizó como elementos de repetición, «células» (del latín cellulae, celdillas). Pero Hooke sólo pudo observar células muertas por lo que no pudo describir las estructuras de su interior.[10]
- Década de 1670: Anton van Leeuwenhoek,
- observó diversas células eucariotas (como protozoos y espermatozoides)
- y procariotas (bacterias).
- 1745: John Needham
- describió la presencia de «animálculos» o «infusorios»; se trataba de organismos unicelulares.

Robert
Hooke bajo su microscopio y tal como aparece publicado en Micrographia.
- Década
- de 1830: Theodor Schwann estudió
- la célula animal;
- junto con Matthias Schleiden postularon que las células son las unidades elementales en la formación de las plantas y animales, y que son la base fundamental del proceso vital.
- 1831: Robert Brown describió el núcleo celular.
- 1839: Purkinje observó
- el citoplasma celular.
- 1850: Rudolf Virchow postuló que todas las células provienen de otras células.
- 1857: Kölliker identificó las mitocondrias.
- 1860: Pasteur realizó multitud de estudios sobre el metabolismo de levaduras y sobre la asepsia.
- 1880: August Weismann descubrió que las células actuales comparten similitud estructural y molecular con células de tiempos remotos.
- 1931: Ernst Ruska construyó el primer microscopio electrónico de
- transmisión en la Universidad de Berlín
- . Cuatro años más tarde, obtuvo un poder de resolución doble a la del microscopio óptico.
- 1981: Lynn Margulis
Definición
Por tanto, podemos definir a la célula como la unidad morfológica y funcional de todo ser vivo. De hecho, la célula es el elemento de menor tamaño que puede considerarse vivo. Como tal posee una membrana de fosfolípidos con permeabilidad selectiva que mantiene un medio interno altamente ordenado y diferenciado del medio externo en cuanto a su composición, sujeta a control homeostático, la cual consiste en biomoléculas y algunos metales y electrolitos. La estructura se automantiene activamente mediante el metabolismo, asegurándose la coordinación de todos los elementos celulares y su perpetuación por replicación a través de un genoma codificado por ácidos nucleicos. La parte de la biología que se ocupa de ella es la citología.Características
Las células, como sistemas termodinámicos complejos, poseen una serie de elementos estructurales y funcionales comunes que posibilitan su supervivencia; no obstante, los distintos tipos celulares presentan modificaciones de estas características comunes que permiten su especialización funcional y, por ello, la ganancia de complejidad.[14] De este modo, las células permanecen altamente organizadas a costa de incrementar la entropía del entorno, uno de los requisitos de la vida.[15]Características estructurales

- Individualidad: Todas las células están rodeadas de una envoltura (que puede ser una bicapa lipídica desnuda, en células animales; una pared de polisacárido, en hongos y vegetales; una membrana externa y otros elementos que definen una pared compleja, en bacterias Gram negativas; una pared de peptidoglicano, en bacterias Gram positivas; o una pared de variada composición, en arqueas)[8] que las separa y comunica con el exterior, que controla los movimientos celulares y que mantiene el potencial de membrana.
- Contienen un medio interno acuoso, el citosol, que forma la mayor parte del volumen celular y en el que están inmersos los orgánulos celulares.
- Poseen material genético en forma de ADN, el material hereditario de los genes y que contiene las instrucciones para el funcionamiento celular, así como ARN, a fin de que el primero se exprese.[16]
- Tienen enzimas y otras proteínas, que sustentan, junto con otras biomoléculas, un metabolismo activo.
Características funcionales

- Nutrición. Las células toman sustancias del medio, las transforman de una forma a otra, liberan energía y eliminan productos de desecho, mediante el metabolismo.
- Crecimiento y multiplicación. Las células son capaces de dirigir su propia síntesis. A consecuencia de los procesos nutricionales, una célula crece y se divide, formando dos células, en una célula idéntica a la célula original, mediante la división celular.
- Diferenciación. Muchas células pueden sufrir cambios de forma o función en un proceso llamado diferenciación celular. Cuando una célula se diferencia, se forman algunas sustancias o estructuras que no estaban previamente formadas y otras que lo estaban dejan de formarse. La diferenciación es a menudo parte del ciclo celular en que las células forman estructuras especializadas relacionadas con la reproducción, la dispersión o la supervivencia.
- Señalización. Las células responden a estímulos químicos y físicos tanto del medio externo como de su interior y, en el caso de células móviles, hacia determinados estímulos ambientales o en dirección opuesta mediante un proceso que se denomina quimiotaxis. Además, frecuentemente las células pueden interaccionar o comunicar con otras células, generalmente por medio de señales o mensajeros químicos, como hormonas, neurotransmisores, factores de crecimiento... en seres pluricelulares en complicados procesos de comunicación celular y transducción de señales.
- Evolución. A diferencia de las estructuras inanimadas, los organismos unicelulares y pluricelulares evolucionan. Esto significa que hay cambios hereditarios (que ocurren a baja frecuencia en todas las células de modo regular) que pueden influir en la adaptación global de la célula o del organismo superior de modo positivo o negativo. El resultado de la evolución es la selección de aquellos organismos mejor adaptados a vivir en un medio particular.
Tamaño, forma y función

En cuanto al tamaño, la mayoría de las células son microscópicas, es decir, no son observables a simple vista. A pesar de ser muy pequeñas (un milímetro cúbico de sangre puede contener unos cinco millones de células),[14] el tamaño de las células es extremadamente variable. La célula más pequeña observada, en condiciones normales, corresponde a Mycoplasma genitalium, de 0,2 μm, encontrándose cerca del límite teórico de 0,17 μm.[21] Existen bacterias con 1 y 2 μm de longitud. Las células humanas son muy variables: hematíes de 7 micras, hepatocitos con 20 micras, espermatozoides de 53 μm, óvulos de 150 μm e, incluso, algunas neuronas de en torno a un metro. En las células vegetales los granos de polen pueden llegar a medir de 200 a 300 μm y algunos huevos de aves pueden alcanzar entre 1 (codorniz) y 7 cm (avestruz) de diámetro. Para la viabilidad de la célula y su correcto funcionamiento siempre se debe tener en cuenta la relación superficie-volumen.[15] Puede aumentar considerablemente el volumen de la célula y no así su superficie de intercambio de membrana lo que dificultaría el nivel y regulación de los intercambios de sustancias vitales para la célula.
Respecto de su forma, las células presentan una gran variabilidad, e, incluso, algunas no la poseen bien definida o permanente. Pueden ser: fusiformes (forma de huso), estrelladas, prismáticas, aplanadas, elípticas, globosas o redondeadas, etc. Algunas tienen una pared rígida y otras no, lo que les permite deformar la membrana y emitir prolongaciones citoplasmáticas (pseudópodos) para desplazarse o conseguir alimento. Hay células libres que no muestran esas estructuras de desplazamiento pero poseen cilios o flagelos, que son estructuras derivadas de un orgánulo celular (el centrosoma) que dota a estas células de movimiento.[2] De este modo, existen multitud de tipos celulares, relacionados con la función que desempeñan; por ejemplo:
- Células contráctiles que suelen ser alargadas, como las fibras musculares.
- Células con finas prolongaciones, como las neuronas que transmiten el impulso nervioso.
- Células con microvellosidades o con pliegues, como las del intestino para ampliar la superficie de contacto y de intercambio de sustancias.
- Células cúbicas, prismáticas o aplanadas como las epiteliales que recubren superficies como las losas de un pavimento.
Estudio de las células
Los biólogos utilizan diversos instrumentos para lograr el conocimiento de las células. Obtienen información de sus formas, tamaños y componentes, que les sirve para comprender además las funciones que en ellas se realizan. Desde las primeras observaciones de células, hace más de 300 años, hasta la época actual, las técnicas y los aparatos se han ido perfeccionando, originándose una rama más de la Biología: la Microscopía. Dado el pequeño tamaño de la gran mayoría de las células, el uso del microscopio es de enorme valor en la investigación biológica. En la actualidad, los biólogos utilizan dos tipos básicos de microscopio: los ópticos y los electrónicos.La célula procariota
Las células procariotas son pequeñas y menos complejas que las eucariotas. Contienen ribosomas pero carecen de sistemas de endomembranas (esto es, orgánulos delimitados por membranas biológicas, como puede ser el núcleo celular). Por ello poseen el material genético en el citosol. Sin embargo, existen excepciones: algunas bacterias fotosintéticas poseen sistemas de membranas internos.[22] También en el Filo Planctomycetes existen organismos como Pirellula que rodean su material genético mediante una membrana intracitoplasmática y Gemmata obscuriglobus que lo rodea con doble membrana. Esta última posee además otros compartimentos internos de membrana, posiblemente conectados con la membrana externa del nucleoide y con la membrana nuclear, que no posee peptidoglucano.[23] [24] [25]Por lo general podría decirse que los procariotas carecen de citoesqueleto. Sin embargo se ha observado que algunas bacterias, como Bacillus subtilis, poseen proteínas tales como MreB y mbl que actúan de un modo similar a la actina y son importantes en la morfología celular.[26] Fusinita van den Ent, en Nature, va más allá, afirmando que los citoesqueletos de actina y tubulina tienen origen procariótico.[27]
De gran diversidad, los procariotas sustentan un metabolismo extraordinariamente complejo, en algunos casos exclusivo de ciertos taxa, como algunos grupos de bacterias, lo que incide en su versatilidad ecológica.[12] Los procariotas se clasifican, según Carl Woese, en arqueas y bacterias.[28]
Arqueas

Las arqueas, al igual que las bacterias, no tienen membranas internas que delimiten orgánulos. Como todos los organismos presentan ribosomas, pero a diferencia de los encontrados en las bacterias que son sensibles a ciertos agentes antimicrobianos, los de las arqueas, más cercanos a los eucariotas, no lo son. La membrana celular tiene una estructura similar a la de las demás células, pero su composición química es única, con enlaces tipo éter en sus lípidos.[30] Casi todas las arqueas poseen una pared celular (algunos Thermoplasma son la excepción) de composición característica, por ejemplo, no contienen peptidoglicano (mureína), propio de bacterias. No obstante pueden clasificarse bajo la tinción de Gram, de vital importancia en la taxonomía de bacterias; sin embargo, en arqueas, poseedoras de una estructura de pared en absoluto común a la bacteriana, dicha tinción es aplicable pero carece de valor taxonómico. El orden Methanobacteriales tiene una capa de pseudomureína, que provoca que dichas arqueas respondan como positivas a la tinción de Gram.[31] [32] [33]
Como en casi todos los procariotas, las células de las arqueas carecen de núcleo, y presentan un sólo cromosoma circular. Existen elementos extracromosómicos, tales como plásmidos. Sus genomas son de pequeño tamaño, sobre 2-4 millones de pares de bases. También es característica la presencia de ARN polimerasas de constitución compleja y un gran número de nucleótidos modificados en los ácidos ribonucleicos ribosomales. Por otra parte, su ADN se empaqueta en forma de nucleosomas, como en los eucariotas, gracias a proteínas semejantes a las histonas y algunos genes poseen intrones.[34] Pueden reproducirse por fisión binaria o múltiple, fragmentación o gemación.
Bacterias

Poseen una membrana celular compuesta de lípidos, en forma de una bicapa y sobre ella se encuentra una cubierta en la que existe un polisacárido complejo denominado peptidoglicano; dependiendo de su estructura y subsecuente su respuesta a la tinción de Gram, se clasifica a las bacterias en Gram positivas y Gram negativas. El espacio comprendido entre la membrana celular y la pared celular (o la membrana externa, si esta existe) se denomina espacio periplásmico. Algunas bacterias presentan una cápsula. Otras son capaces de generar endosporas (estadios latentes capaces de resistir condiciones extremas) en algún momento de su ciclo vital. Entre las formaciones exteriores propias de la célula bacteriana destacan los flagelos (de estructura completamente distinta a la de los flagelos eucariotas) y los pili (estructuras de adherencia y relacionadas con la parasexualidad).[8]
La mayoría de las bacterias disponen de un único cromosoma circular y suelen poseer elementos genéticos adicionales, como distintos tipos de plásmidos. Su reproducción, binaria y muy eficiente en el tiempo, permite la rápida expansión de sus poblaciones, generándose un gran número de células que son virtualmente clones, esto es, idénticas entre sí.[34]
La célula eucariota
Las células eucariotas son el exponente de la complejidad celular actual.[14] Presentan una estructura básica relativamente estable caracterizada por la presencia de distintos tipos de orgánulos intracitoplasmáticos especializados, entre los cuales destaca el núcleo, que alberga el material genético. Especialmente en los organismos pluricelulares, las células pueden alcanzar un alto grado de especialización. Dicha especialización o diferenciación es tal que, en algunos casos, compromete la propia viabilidad del tipo celular en aislamiento. Así, por ejemplo, las neuronas dependen para su supervivencia de las células gliales.[12] Por otro lado, la estructura de la célula varía dependiendo de la situación taxonómica del ser vivo: de este modo, las células vegetales difieren de las animales, así como de las de los hongos. Por ejemplo, las células animales carecen de pared celular, son muy variables, no tiene plastos, puede tener vacuolas pero no son muy grandes y presentan centríolos (que son agregados de microtúbulos cilíndricos que contribuyen a la formación de los cilios y los flagelos y facilitan la división celular). Las células de los vegetales, por su lado, presentan una pared celular compuesta principalmente de celulosa), disponen de plastos como cloroplastos (orgánulo capaz de realizar la fotosíntesis), cromoplastos (orgánulos que acumulan pigmentos) o leucoplastos (orgánulos que acumulan el almidón fabricado en la fotosíntesis), poseen vacuolas de gran tamaño que acumulan sustancias de reserva o de desecho producidas por la célula y finalmente cuentan también con plasmodesmos, que son conexiones citoplasmáticas que permiten la circulación directa de las sustancias del citoplasma de una célula a otra, con continuidad de sus membranas plasmáticas.[37]Compartimentos
Las células son entes dinámicos, con un metabolismo celular interno de gran actividad cuya estructura es un flujo entre rutas anastomosadas. Un fenómeno observado en todos los tipos celulares es la compartimentalización, que consiste en una heterogeneidad que da lugar a entornos más o menos definidos (rodeados o no mediante membranas biológicas) en las cuales existe un microentorno que aglutina a los elementos implicados en una ruta biológica.[38] Esta compartimentalización alcanza su máximo exponente en las células eucariotas, las cuales están formadas por diferentes estructuras y orgánulos que desarrollan funciones específicas, lo que supone un método de especialización espacial y temporal.[2] No obstante, células más sencillas, como los procariotas, ya poseen especializaciones semejantes.[39]Membrana plasmática y superficie celular
La composición de la membrana plasmática varía entre células dependiendo de la función o del tejido en la que se encuentre, pero posee elementos comunes. Está compuesta por una doble capa de fosfolípidos, por proteínas unidas no covalentemente a esa bicapa, y por glúcidos unidos covalentemente a lípidos o proteínas. Generalmente, las moléculas más numerosas son las de lípidos; sin embargo, la proteínas, debido a su mayor masa molecular, representan aproximadamente el 50% de la masa de la membrana.[38]Un modelo que explica el funcionamiento de la membrana plasmática es el modelo del mosaico fluido, de J. S. Singer y Garth Nicolson (1972), que desarrolla un concepto de unidad termodinámica basada en las interacciones hidrófobas entre moléculas y otro tipo de enlaces no covalentes.[40]

Sobre la bicapa lipídica, independientemente de la presencia o no de una pared celular, existe una matriz que puede variar, de poco conspicua, como en los epitelios, a muy extensa, como en el tejido conjuntivo. Dicha matriz, denominada glucocalix (glicocáliz), rica en líquido tisular, glucoproteínas, proteoglicanos y fibras, también interviene en la generación de estructuras y funciones emergentes, derivadas de las interacciones célula-célula.[12]
Estructura y expresión génica

Dicho material genético se encuentra inmerso en una actividad continua de regulación de la expresión génica; las ARN polimerasas transcriben ARN mensajero continuamente, que, exportado al citosol, es traducido a proteína, de acuerdo a las necesidades fisiológicas. Asimismo, dependiendo del momento del ciclo celular, dicho ADN puede entrar en replicación, como paso previo a la mitosis.[34] No obstante, las células eucarióticas poseen material genético extranuclear: concretamente, en mitocondrias y plastos, si los hubiere; estos orgánulos conservan una independencia genética parcial del genoma nuclear.[43] [44]
Síntesis y degradación de macromoléculas
Dentro del citosol, esto es, la matriz acuosa que alberga a los orgánulos y demás estructuras celulares, se encuentran inmersos multitud de tipos de maquinaria de metabolismo celular: orgánulos, inclusiones, elementos del citoesqueleto, enzimas... De hecho, estas últimas corresponden al 20% de las enzimas totales de la célula.[12]

- Ribosoma: Los ribosomas, visibles al microscopio electrónico como partículas esféricas,[45] son complejos supramoleculares encargados de ensamblar proteínas a partir de la información genética que les llega del ADN transcrita en forma de ARN mensajero. Elaborados en el núcleo, desempeñan su función de síntesis de proteínas en el citoplasma. Están formados por ARN ribosómico y por diversos tipos de proteínas. Estructuralmente, tienen dos subunidades. En las células, estos orgánulos aparecen en diferentes estados de disociación. Cuando están completos, pueden estar aislados o formando grupos (polisomas). También pueden aparecer asociados al retículo endoplasmático rugoso o a la envoltura nuclear.[34]
- Retículo endoplasmático: El retículo endoplasmático es orgánulo vesicular interconectado que forma cisternas, tubos aplanados y sáculos comunicados entre sí. Intervienen en funciones relacionadas con la síntesis proteica, glicosilación de proteínas, metabolismo de lípidos y algunos esteroides, detoxificación, así como el tráfico de vesículas. En células especializadas, como las miofibrillas o células musculares, se diferencia en el retículo sarcoplásmico, orgánulo decisivo para que se produzca la contracción muscular.[14]
- Aparato de Golgi: El aparato de Golgi es un orgánulo formado por apilamientos de sáculos denominados dictiosomas, si bien, como ente dinámico, estos pueden interpretarse como estructuras puntuales fruto de la coalescencia de vesículas.[46] [47] Recibe las vesículas del retículo endoplasmático rugoso que han de seguir siendo procesadas. Dentro de las funciones que posee el aparato de Golgi se encuentran la glicosilación de proteínas, selección, destinación, glicosilación de lípidos y la síntesis de polisacáridos de la matriz extracelular. Posee tres compartimientos; uno proximal al retículo endoplasmático, denominado «compartimento cis», donde se produce la fosforilación de las manosas de las enzimas que han de dirigirse al lisosoma; el «compartimento intermedio», con abundantes manosidasas y N-acetil-glucosamina transferasas; y el «compartimento o red trans», el más distal, donde se transfieren residuos de galactosa y ácido siálico, y del que emergen las vesículas con los diversos destinos celulares.[12]
- Lisosoma: Los lisosomas son orgánulos que albergan multitud de enzimas hidrolíticas. De morfología muy variable, no se ha demostrado su existencia en células vegetales.[12] Una característica que agrupa a todos los lisosomas es la posesión de hidrolasas ácidas: proteasas, nucleasas, glucosidasas, lisozima, arilsulfatasas, lipasas, fosfolipasas y fosfatasas. Procede de la fusión de vesículas procedentes del aparato de Golgi, que, a su vez, se fusionan en un tipo de orgánulo denominado endosoma temprano, el cual, al acidificarse y ganar en enzimas hidrolíticos, pasa a convertirse en el lisosoma funcional. Sus funciones abarcan desde la degradación de macromoléculas endógenas o procedentes de la fagocitosis a la intervención en procesos de apoptosis.[48]

- Vacuola vegetal: Las vacuolas vegetales, numerosas y pequeñas en células meristemáticas y escasas y grandes en células diferenciadas, son orgánulos exclusivos de los representantes del mundo vegetal. Inmersas en el citosol, están delimitadas por el tonoplasto, una membrana lipídica. Sus funciones son: facilitar el intercambio con el medio externo, mantener la turgencia celular, la digestión celular y la acumulación de sustancias de reserva y subproductos del metabolismo.[37]
- Inclusión citoplasmática: Las inclusiones son acúmulos nunca delimitados por membrana de sustancias de diversa índole, tanto en células vegetales como animales. Típicamente se trata de sustancias de reserva que se conservan como acervo metabólico: almidón, glucógeno, triglicéridos, proteínas... aunque también existen de pigmentos.[12]
Conversión energética
El metabolismo celular está basado en la transformación de unas sustancias químicas, denominadas metabolitos, en otras; dichas reacciones químicas transcurren catalizadas mediante enzimas. Si bien buena parte del metabolismo sucede en el citosol, como la glucólisis, existen procesos específicos de orgánulos.[41]
- Mitocondria: Las mitocondrias son orgánulos de aspecto, número y tamaño variable que intervienen en el ciclo de Krebs, fosforilación oxidativa y en la cadena de transporte de electrones de la respiración. Presentan una doble membrana, externa e interna, que dejan entre ellas un espacio perimitocondrial; la membrana interna, plegada en crestas hacia el interior de la matriz mitocondrial, posee una gran superficie. En su interior posee generalmente una sola molécula de ADN, el genoma mitocondrial, típicamente circular, así como ribosomas más semejantes a los bacterianos que a los eucariotas.[12] Según la teoría endosimbiótica, se asume que la primera protomitocondria era un tipo de proteobacteria.[49]

- Cloroplasto: Los cloroplastos son los orgánulos celulares que en los organismos eucariotas fotosintéticos se ocupan de la fotosíntesis. Están limitados por una envoltura formada por dos membranas concéntricas y contienen vesículas, los tilacoides, donde se encuentran organizados los pigmentos y demás moléculas implicadas en la conversión de la energía luminosa en energía química. Además de esta función, los plastidios intervienen en el metabolismo intermedio, produciendo energía y poder reductor, sintetizando bases púricas y pirimidínicas, algunos aminoácidos y todos los ácidos grasos. Además, en su interior es común la acumulación de sustancias de reserva, como el almidón.[12] Se considera que poseen analogía con las cianobacterias.[50]

- Peroxisoma: Los peroxisomas son orgánulos muy comunes en forma de vesículas que contienen abundantes enzimas de tipo oxidasa y catalasa; de tan abundantes, es común que cristalicen en su interior. Estas enzimas cumplen funciones de detoxificación celular. Otras funciones de los peroxisomas son: las oxidaciones flavínicas generales, el catabolismo de las purinas, la beta-oxidación de los ácidos grasos, el ciclo del glioxilato, el metabolismo del ácido glicólico y la detoxificación en general.[12] Se forman de vesículas procedentes del retículo endoplasmático.[51]
Citoesqueleto
Las células poseen un andamiaje que permite el mantenimiento de su forma y estructura, pero más aún, este es un sistema dinámico que interactúa con el resto de componentes celulares generando un alto grado de orden interno. Dicho andamiaje está formado por una serie de proteínas que se agrupan dando lugar a estructuras filamentosas que, mediante otras proteínas, interactúan entre ellas dando lugar a una especie de retículo. El mencionado andamiaje recibe el nombre de citoesqueleto, y sus elementos mayoritarios son: los microtúbulos, los microfilamentos y los filamentos intermedios.[2] [nota 2] [52] [53]- Microfilamentos: Los microfilamentos o filamentos de actina están formados por una proteína globular, la actina, que puede polimerizar dando lugar a estructuras filiformes. Dicha actina se expresa en todas las células del cuerpo y especialmente en las musculares ya que está implicada en la contracción muscular, por interacción con la miosina. Además, posee lugares de unión a ATP, lo que dota a sus filamentos de polaridad.[54] Puede encontrarse en forma libre o polimerizarse en microfilamentos, que son esenciales para funciones celulares tan importantes como la movilidad y la contracción de la célula durante la división celular.[46]

- Microtúbulos: Los microtúbulos son estructuras tubulares de 25 nm de diámetro exterior y unos 12 nm de diámetro interior, con longitudes que varían entre unos pocos nanómetros a micrómetros, que se originan en los centros organizadores de microtúbulos y que se extienden a lo largo de todo el citoplasma. Se hallan en las células eucariotas y están formadas por la polimerización de un dímero de dos proteínas globulares, la alfa y la beta tubulina. Las tubulinas poseen capacidad de unir GTP.[2] [46] Los microtúbulos intervienen en diversos procesos celulares que involucran desplazamiento de vesículas de secreción, movimiento de orgánulos, transporte intracelular de sustancias, así como en la división celular (mitosis y meiosis) y que, junto con los microfilamentos y los filamentos intermedios, forman el citoesqueleto. Además, constituyen la estructura interna de los cilios y los flagelos.[2] [46]
- Filamentos intermedios: Los filamentos intermedios son componentes del citoesqueleto. Formados por agrupaciones de proteínas fibrosas, su nombre deriva de su diámetro, de 10 nm, menor que el de los microtúbulos, de 24 nm, pero mayor que el de los microfilamentos, de 7 nm. Son ubicuos en las células animales, y no existen en plantas ni hongos. Forman un grupo heterogéneo, clasificado en cinco familias: las queratinas, en células epiteliales; los neurofilamentos, en neuronas; los gliofilamentos, en células gliales; la desmina, en músculo liso y estriado; y la vimentina, en células derivadas del mesénquima.[12]

- Centríolos: Los centríolos son una pareja de estructuras que forman parte del citoesqueleto de células animales. Semejantes a cilindros huecos, están rodeados de un material proteico denso llamado material pericentriolar; todos ellos forman el centrosoma o centro organizador de microtúbulos que permiten la polimerización de microtúbulos de dímeros de tubulina que forman parte del citoesqueleto. Los centríolos se posicionan perpendicularmente entre sí. Sus funciones son participar en la mitosis, durante la cual generan el huso acromático, y en la citocinesis,[55] así como, se postula, intervenir en la nucleación de microtúbulos.[56] [57]
- Cilios y flagelos: Se trata de especializaciones de la superficie celular con motilidad; con una estructura basada en agrupaciones de microtúbulos, ambos se diferencian en la mayor longitud y menor número de los flagelos, y en la mayor variabilidad de la estructura molecular de estos últimos.[12]
Ciclo vital

- El estado de no división o interfase. La célula realiza sus funciones específicas y, si está destinada a avanzar a la división celular, comienza por realizar la duplicación de su ADN.
- El estado de división, llamado fase M, situación que comprende la mitosis y citocinesis. En algunas células la citocinesis no se produce, obteniéndose como resultado de la división una masa celular plurinucleada denominada plasmodio.[nota 3]
- La interfase consta de tres estadios claramente definidos.[2]
[46]
- Fase G1: es la primera fase del ciclo celular, en la que existe crecimiento celular con síntesis de proteínas y de ARN. Es el período que trascurre entre el fin de una mitosis y el inicio de la síntesis de ADN. En él la célula dobla su tamaño y masa debido a la continua síntesis de todos sus componentes, como resultado de la expresión de los genes que codifican las proteínas responsables de su fenotipo particular.
- Fase S: es la segunda fase del ciclo, en la que se produce la replicación o síntesis del ADN. Como resultado cada cromosoma se duplica y queda formado por dos cromátidas idénticas. Con la duplicación del ADN, el núcleo contiene el doble de proteínas nucleares y de ADN que al principio.
- Fase G2: es la segunda fase de crecimiento del ciclo celular en la que continúa la síntesis de proteínas y ARN. Al final de este período se observa al microscopio cambios en la estructura celular, que indican el principio de la división celular. Termina cuando los cromosomas empiezan a condensarse al inicio de la mitosis.
- La fase M es la fase de la división celular en la cual una célula progenitora se divide en dos células hijas hijas idénticas entre sí y a la madre. Esta fase incluye la mitosis, a su vez dividida en: profase, metafase, anafase, telofase; y la citocinesis, que se inicia ya en la telofase mitótica.
Origen
La aparición de la vida, y, por ello, de la célula, probablemente se inició gracias a la transformación de moléculas inorgánicas en orgánicas bajo unas condiciones ambientales adecuadas, produciéndose más adelante la interacción de estas biomoléculas generando entes de mayor complejidad. El experimento de Miller y Urey, realizado en 1953, demostró que una mezcla de compuestos orgánicos sencillos puede transformarse en algunos aminoácidos, glúcidos y lípidos (componentes todos ellos de la materia viva) bajo unas condiciones ambientales que simulan las presentes hipotéticamente en la Tierra primigenia (en torno al eón Arcaico).[60]Se postula que dichos componentes orgánicos se agruparon generando estructuras complejas, los coacervados de Oparin, aún acelulares que, en cuanto alcanzaron la capacidad de autoorganizarse y perpetuarse, dieron lugar a un tipo de célula primitiva, el progenote de Carl Woese, antecesor de los tipos celulares actuales.[28] Una vez se diversificó este grupo celular, dando lugar a las variantes procariotas, arqueas y bacterias, pudieron aparecer nuevos tipos de células, más complejos, por endosimbiosis, esto es, captación permanente de unos tipos celulares en otros sin una pérdida total de autonomía de aquellos.[61] De este modo, algunos autores describen un modelo en el cual la primera célula eucariota surgió por introducción de una arquea en el interior de una bacteria, dando lugar esta primera a un primitivo núcleo celular.[62] No obstante, la imposibilidad de que una bacteria pueda efectuar una fagocitosis y, por ello, captar a otro tipo de célula, dio lugar a otra hipótesis, que sugiere que fue una célula denominada cronocito la que fagocitó a una bacteria y a una arquea, dando lugar al primer organismo eucariota. De este modo, y mediante un análisis de secuencias a nivel genómico de organismos modelo eucariotas, se ha conseguido describir a este cronocito original como un organismo con citoesqueleto y membrana plasmática, lo cual sustenta su capacidad fagocítica, y cuyo material genético era el ARN, lo que puede explicar, si la arquea fagocitada lo poseía en el ADN, la separación espacial en los eucariotas actuales entre la transcripción (nuclear), y la traducción (citoplasmática).[63]
Una dificultad adicional es el hecho de que no se han encontrado organismos eucariotas primitivamente amitocondriados como exige la hipótesis endosimbionte. Además, el equipo de María Rivera, de la Universidad de California, comparando genomas completos de todos los dominios de la vida ha encontrado evidencias de que los eucariotas contienen dos genomas diferentes, uno más semejante a bacterias y otro a arqueas, apuntando en este último caso semejanzas a los metanógenos, en particular en el caso de las histonas.[64] [65] Esto llevó a Bill Martin y Miklós Müller a plantear la hipótesis de que la célula eucariota surgiera no por endosimbiosis, sino por fusión quimérica y acoplamiento metabólico de un metanógeno y una α-proteobacteria simbiontes a través del hidrógeno (hipótesis del hidrógeno).[66] Esta hipótesis atrae hoy en día posiciones muy encotradas, con detractores como Christian de Duve.[67]
Harold Morowitz, un físico de la Universidad Yale, ha calculado que las probabilidades de obtener la bacteria viva más sencilla mediante cambios al azar es de 1 sobre 1 seguido por 100.000.000.000 de ceros. “Este número es tan grande —dijo Robert Shapiro— que para escribirlo en forma convencional necesitaríamos varios centenares de miles de libros en blanco.” Presenta la acusación de que los científicos que han abrazado la evolución química de la vida pasan por alto la evidencia aumentante y “han optado por aceptarla como verdad que no puede ser cuestionada, consagrándola así como mitología”.[68]
El microscopio (de micr
o-, μικρο, pequeño, y scopio, σκοπεω, observar) es un instrumento que permite observar objetos que son demasiado pequeños para ser vistos a simple vista. El tipo más común y el primero que se inventó es el microscopio óptico. Se trata de un instrumento óptico que contiene dos o más lentes que permiten obtener una imagen aumentada del objeto y que funciona por refracción. La ciencia que investiga los objetos pequeños utilizando este instrumento se llama microscopía.

Partes del microscopio
- Lente ocular.Texto grande
- Revólver.
- Platina.
- Lentes óptico.
- Diafragma.
- Pinzas de la platina.
- Base o pie.
- Lámpara.
- Tornillo micrométrico(ajuste fino).
- Tornillo macrométrico(de cremallera).
- Brazo o columna.
- Tubo ocular.

fabricado hacia 1751 por Magny.
Proviene del laboratorio del duque de
Chaulnes y pertenece al Museo de Artes y +Oficios, París
fue inventado hacia los años 1610, por Galileo según los italianos
, o por Zacharias Janssen en 1590, en opinión de los holandeses. En 1665 aparece en la obra de William Harvey sobre la circulación sanguínea al observar al microscopio los capilares sanguíneos y Robert Hooke publica su obra Micrographia.
En 1665 Robert Hooke observó con un microscopio un delgado corte
de corcho y notó que el material era poroso,
en su conjunto, formaban cavidades poco profundas a modo de celditas a las que llamó células. Se trataba de la primera observación de células muertas. Unos años más tarde, Marcello Malpighi, anatomista y biólogo italiano, observó células vivas. Fue el primero en estudiar tejidos vivos al microscopio.
A mediados del siglo XVII
un holandés, Anton van Leeuwenhoek, utilizando microscopios simples de fabricación propia, describió por primera vez protozoos, bacterias, espermatozoides y glóbulos rojos. El microscopista Leeuwenhoek, sin ninguna preparación científica, puede considerarse el fundador de la bacteriología. Tallaba él mismo sus lupas, sobre pequeñas esferas de cristal, cuyos diámetros no alcanzaban el milímetro (su campo de visión era muy limitado, de décimas de milímetro). Con estas pequeñas distancias focales alcanzaba los 275 aumentos. Observó los glóbulos de la sangre, las bacterias y los protozoos; examinó por primera vez los glóbulos rojos y descubrió que el semen contiene espermatozoides. Durante su vida no reveló sus métodos secretos y a su muerte, en 1723, 26 de sus aparatos fueron cedidos a la Royal Society de Londres.
Durante el siglo XVIII continuó el progreso y se lograron objetivos acromáticos por asociación de Chris Neros y Flint Crown obtenidos en 1740 por H. M. Hall y mejorados por John Dollond. De esta época son los estudios efectuados por Isaac Newton y Leonhard Euler. En el siglo XIX, al descubrirse que la dispersión y la refracción se podían modificar con combinaciones adecuadas de dos o más medios ópticos, se lanzan al mercado objetivos acromáticos excelentes.
Durante el siglo XVIII el microscopio tuvo diversos adelantos mecánicos que aumentaron su estabilidad y su facilidad de uso, aunque no se desarrollaron por el momento mejoras ópticas. Las mejoras más importantes de la óptica surgieron en 1877, cuando Ernst Abbe publicó su teoría del microscopio y, por encargo de Carl Zeiss, mejoró la microscopía de inmersión sustituyendo el agua por aceite de cedro, lo que permite obtener aumentos de 2000. A principios de los años 1930 se había alcanzado el límite teórico para los microscopios ópticos, no consiguiendo estos aumentos superiores a 500X o 1,000X. Sin embargo, existía un deseo científico de observar los detalles de estructuras celulares (núcleo, mitocondria, etc.).
El microscopio electrónico de transmisión (TEM) fue el primer tipo de microscopio electrónico desarrollado. Utiliza un haz de electrones en lugar de luz para enfocar la muestra consiguiendo aumentos de 100.000X. Fue desarrollado por Max Knoll y Ernst Ruska en Alemania en 1931. Posteriormente, en 1942 se desarrolla el microscopio electrónico de barrido (SEM).
Tipos de microscopios

- Microscopio óptico
- Microscopio simple
- Microscopio compuesto
- Microscopio de luz ultravioleta
- Microscopio de fluorescencia
- Microscopio petrográfico
- Microscopio en campo oscuro
- Microscopio de contraste de fase
- Microscopio de luz polarizada
- Microscopio confocal
- Microscopio electrónico
- Microscopio electrónico de transmisión
- Microscopio electrónico de barrido
- Microscopio de iones en campo
- Microscopio de sonda de barrido
- Microscopio de efecto túnel
- Microscopio de fuerza atómica
- Microscopio virtual
- Microscopio de antimateria
- Microscopio reflector
- Microscopio telegramatico
- Microscopio nuclear
Átomo

El átomo
es la unidad de materia más pequeña
de un elemento químico que mantiene su identidad o sus propiedades, y que no es posible dividir mediante procesos químicos. Está compuesto por un núcleo atómico, en el que se concentra casi toda su masa, rodeado de una nube de electrones. El núcleo está formado por protones, con carga positiva, y neutrones, eléctricamente neutros.[nota 1] Los electrones, cargados negativamente, permanecen ligados a este mediante la fuerza electromagnética.
Los átomos se clasifican de acuerdo
al número de protones y neutrones que contenga su núcleo. El número de protones o número atómico determina su elemento químico, y el número de neutrones determina su isótopo. Un átomo con el mismo número de protones que de electrones es eléctricamente neutro. Si por el contrario posee un exceso de protones o de electrones, su carga neta es positiva o negativa, y se denomina ion.
El nombre átomo
proviene del latín atomum, y éste del griego ἄτομον, sin partes; también, se deriva de «a» (no) y «tomo» (divisible); no divisible.[1] El concepto de átomo como bloque básico e indivisible que compone la materia del universo fue postulado por la escuela atomista en la Antigua Grecia. Sin embargo, su existencia no quedó demostrada hasta el siglo XIX. Con el desarrollo de la física nuclear en el siglo XX se comprobó que el átomo puede subdividirse en partículas más pequeñas.[2] [3]
Los átomos son objetos muy pequeños
con masas igualmente minúsculas: su diámetro y masa son del orden de la billonésima parte de un metro y cuatrillonésima parte de un gramo. Solo pueden ser observados mediante instrumentos especiales tales como un microscopio de efecto túnel. Más de un 99,94% de la masa del átomo está concentrada en su núcleo, en general repartida de manera aproximadamente equitativa entre protones y neutrones. El núcleo de un átomo puede ser inestable y sufrir una transmutación mediante desintegración radioactiva. Los electrones en la nube del átomo están repartidos en distintos niveles de energía u orbitales, y determinan las propiedades químicas del mismo. Las transiciones entre los distintos niveles dan lugar a la emisión o absorción de radiación electromagnética en forma de fotones, y son la base de la espectroscopía.
es la unidad de materia más pequeña
de un elemento químico que mantiene su identidad o sus propiedades, y que no es posible dividir mediante procesos químicos. Está compuesto por un núcleo atómico, en el que se concentra casi toda su masa, rodeado de una nube de electrones. El núcleo está formado por protones, con carga positiva, y neutrones, eléctricamente neutros.[nota 1] Los electrones, cargados negativamente, permanecen ligados a este mediante la fuerza electromagnética.
Los átomos se clasifican de acuerdo
al número de protones y neutrones que contenga su núcleo. El número de protones o número atómico determina su elemento químico, y el número de neutrones determina su isótopo. Un átomo con el mismo número de protones que de electrones es eléctricamente neutro. Si por el contrario posee un exceso de protones o de electrones, su carga neta es positiva o negativa, y se denomina ion.
El nombre átomo
proviene del latín atomum, y éste del griego ἄτομον, sin partes; también, se deriva de «a» (no) y «tomo» (divisible); no divisible.[1] El concepto de átomo como bloque básico e indivisible que compone la materia del universo fue postulado por la escuela atomista en la Antigua Grecia. Sin embargo, su existencia no quedó demostrada hasta el siglo XIX. Con el desarrollo de la física nuclear en el siglo XX se comprobó que el átomo puede subdividirse en partículas más pequeñas.[2] [3]
Los átomos son objetos muy pequeños
con masas igualmente minúsculas: su diámetro y masa son del orden de la billonésima parte de un metro y cuatrillonésima parte de un gramo. Solo pueden ser observados mediante instrumentos especiales tales como un microscopio de efecto túnel. Más de un 99,94% de la masa del átomo está concentrada en su núcleo, en general repartida de manera aproximadamente equitativa entre protones y neutrones. El núcleo de un átomo puede ser inestable y sufrir una transmutación mediante desintegración radioactiva. Los electrones en la nube del átomo están repartidos en distintos niveles de energía u orbitales, y determinan las propiedades químicas del mismo. Las transiciones entre los distintos niveles dan lugar a la emisión o absorción de radiación electromagnética en forma de fotones, y son la base de la espectroscopía.
A pesar de que átomo significa
‘indivisible’, en realidad está formado por varias partículas subatómicas. El átomo contiene protones, neutrones y electrones, con la excepción del hidrógeno-1, que no contiene neutrones, y del ion hidronio, que no contiene electrones. Los protones y neutrones del átomo se denominan nucleones, por formar parte del núcleo atómico.
El electrón
es la partícula más ligera de cuantas componen el átomo, con una masa de 9,11 · 10−31 kg. Tiene una carga eléctrica negativa cuya magnitud se define como la carga eléctrica elemental, y se ignora si posee subestructura, por lo que se lo considera una partícula elemental. Los protones tienen una masa de 1,67 · 10−27 kg, 1836 veces la del electrón, y una carga positiva opuesta a la de este. Los neutrones tienen un masa de 1,69 · 10−27 kg, 1839 veces la del electrón, y no poseen carga eléctrica. Las masas de ambos nucleones son ligeramente inferiores dentro del núcleo, debido a la energía potencial del mismo; y sus tamaños son similares, con un radio del orden de 8 · 10-16 m o 0,8 femtómetros (fm).[4]
El protón y el neutrón
no son partículas elementales, sino que constituyen un estado ligado de quarks u y d, partículas fundamentales recogidas en el modelo estándar de la física de partículas, con cargas eléctricas iguales a +2/3 y −1/3 respectivamente, respecto de la carga elemental. Un protón contiene dos quarks u y un quark d, mientras que el neutrón contiene dos d y un u, en consonancia con la carga de ambos. Los quarks se mantienen unidos mediante la fuerza nuclear fuerte, mediada por gluones —del mismo modo que la fuerza electromagnética está mediada por fotones—. Además de estas, existen otras partículas subatómicas en el modelo estándar: más tipos de quarks, leptones cargados (similares al electrón), etc.
se encuentran ligados en el núcleo atómico, la parte central del mismo. El volumen del núcleo es aproximadamente proporcional al número total de nucleones, el número másico A,[5] lo cual es mucho menor que el tamaño del átomo, cuyo radio es del orden de 105 fm o 1 ångström (Å). Los nucleones se mantienen unidos mediante la fuerza nuclear, que es mucho más intensa que la fuerza electromagnética a distancias cortas, lo cual permite vencer la repulsión eléctrica entre los protones.[6]
Los átomos de un
mismo elemento tienen el mismo número de protones, que se denomina
número atómico y se representa por Z. Los átomos de un elemento dado pueden tener distinto número de neutrones: se dice entonces que son isótopos. Ambos números conjuntamente determinan el núclido.
El núcleo atómico puede verse alterado por procesos muy
energéticos en comparación con las reacciones químicas. Los núcleos inestables sufren desintegraciones que pueden cambiar su número de protones y neutrones emitiendo radiación. Un núcleo pesado puede fisionarse en otros más ligeros en una reacción nuclear o espontáneamente. Mediante una cantidad suficiente de energía, dos o más núcleos pueden fusionarse en otro más pesado.
En átomos con
número atómico bajo, los núcleos con una cantidad distinta de protones y neutrones tienden a desintegrarse en núcleos con proporciones más parejas, más estables. Sin embargo, para valores mayores del número atómico, la repulsión mutua de los protones requiere una proporción mayor de neutrones para estabilizar el núcleo.[7]

 Los cinco primeros
Los electrones
Los cinco primeros
Los electrones
en el átomo son atraídos por los protones a través de la fuerza electromagnética. Esta fuerza los atrapa en un pozo de potencial electrostático alrededor del núcleo, lo que hace necesaria una fuente de energía externa para liberarlos. Cuanto más cerca está un electrón del núcleo, mayor es la fuerza atractiva, y mayor por tanto la energía necesaria para que escape.
Los electrones, como
otras partículas, presentan simultáneamente propiedades de partícula puntual y de onda, y tienden a formar un cierto tipo de onda estacionaria alrededor del núcleo, en reposo respecto de este. Cada una de estas ondas está caracterizada por un orbital atómico, una función matemática que describe la probabilidad de encontrar al electrón en cada punto del espacio. El conjunto de estos orbitales es discreto, es decir, puede enumerarse, como es propio en todo sistema cuántico. La nube de electrones es la región ocupada por estas ondas, visualizada como una densidad de carga negativa alrededor del núcleo.
Cada orbital corresponde
a un posible valor de energía para los electrones, que se reparten entre ellos. El principio de exclusión de Pauli prohibe que más de dos electrones se encuentren en el mismo orbital. Pueden ocurrir transiciones entre los distintos niveles de energía: si un electrón absorbe un fotón con energía suficiente, puede saltar a un nivel superior; también desde un nivel más alto puede acabar en un nivel inferior, radiando el resto de la energía en un fotón. Las energías dadas por las diferencias entre los valores de estos niveles son las que se observan en las líneas espectrales del átomo.
En química se utiliza también el mol como unidad de masa. Un mol de átomos de cualquier elemento equivale siempre al mismo número de estos (6,022 · 1023), lo cual implica que un mol de átomos de un elemento con masa atómica de 1 u pesa aproximadamente 1 gramo. En general, un mol de átomos de un cierto elemento pesa de forma aproximada tantos gramos como la masa atómica de dicho elemento.
Los diversos métodos existentes arrojan valores para el radio atómico de entre 0,5 y 5 Å. Dentro de la tabla periódica de los elementos, el tamaño de los átomos tiende a disminuir a lo largo de un periodo —una fila—, para aumentar súbitamente al comienzo de uno nuevo, a medida que los electrones ocupan niveles de energía más altos.[9]
Las dimensiones del átomo son miles de veces más pequeñas que la longitud de onda de la luz (400-700 nm) por lo que estos no pueden ser observados utilizando instrumentos ópticos. En comparación, el grosor de un cabello humano es equivalente a un millón de átomos de carbono. Si una manzana fuera del tamaño de la Tierra, los átomos en ella serían tan grandes como la manzana original.[10]
Cuando un electrón efectúa una transición entre dos estados distintos, absorbe o emite un fotón, cuya energía es precisamente la diferencia entre los dos niveles. La energía de un fotón es proporcional a su frecuencia, así que cada transición se corresponde con una banda estrecha del espectro electromagnético denominada línea espectral.

Un ejemplo de líneas de absorción en un espectro
Cada elemento químico posee un espectro de líneas característico.
Estas se detectan como líneas de emisión en la radiación de los átomos
del mismo. Por el contrario, si se hace pasar radiación con un espectro
de frecuencias continuo a través de estos, los fotones con la energía
adecuada son absorbidos. Cuando los electrones excitados decaen más
tarde, emiten en direcciones aleatorias, por lo que las frecuencias
características se observan como líneas de absorción oscuras. Las medidas espectroscópicas de la intensidad y
anchura de estas líneas permite determinar la composición de una
sustancia.
Algunas líneas espectrales se presentan muy juntas entre sí, tanto que llegaron a confundirse con una sola históricamente, hasta que fue descubierta su subestructura o estructura fina. La causa de este fenómeno se encuentra en las diversas correcciones a considerar en la interacción entre los electrones y el núcleo. Teniendo en cuenta tan solo la fuerza electrostática, ocurre que algunas de las configuraciones electrónicas pueden tener la misma energía aun siendo distintas. El resto de pequeños efectos y fuerzas en el sistema electrón-núcleo rompe esta redundancia o degeneración, dando lugar a la estructura fina. Estos incluyen las correcciones relativistas al movimiento de electrón, la interacción de su momento magnético con el campo eléctrico y con el núcleo, etc.[11]
Además, en presencia de un campo externo los niveles de energía se ven modificados por la interacción del electrón con este, en general produciendo o aumentando la división entre los niveles de energía. Este fenómeno se conoce como efecto Stark en el caso de un campo eléctrico, y efecto Zeeman en el caso de un campo magnético.
Las transiciones de un electrón a un nivel superior ocurren en presencia de radiación electromagnética externa, que provoca la absorción del fotón necesario. Si la frecuencia de dicha radiación es muy alta, el fotón es muy energético y el electrón puede liberarse, en el llamado efecto fotoeléctrico.
Las transiciones a un nivel inferior pueden ocurrir de manera espontánea, emitiendo la energía mediante un fotón saliente; o de manera estimulada, de nuevo en presencia de radiación. En este caso, un fotón «entrante» apropiado provoca que el electrón decaiga a un nivel con una diferencia de energía igual a la del fotón entrante. De este modo, se emite un fotón saliente cuya onda asociada está sincronizada con la del primero, y en la misma dirección. Este fenómeno es la base del láser.
Este tipo de estructura del átomo llevó a Rutherford a proponer su modelo en que los electrones se moverían alrededor del núcleo en órbitas. Este modelo tiene una dificultad proveniente del hecho de que una partícula cargada acelerada, como sería necesario para mantenerse en órbita, radiaría radiación electromagnética, perdiendo energía. Las leyes de Newton, junto con las ecuaciones de Maxwell del electromagnetismo aplicadas al átomo de Rutherford llevan a que en un tiempo del orden de 10−10 s, toda la energía del átomo se habría radiado, con la consiguiente caída de los electrones sobre el núcleo.[13]
Luego en 1811, el físico italiano Amedeo Avogadro, postuló que a una temperatura, presión y volumen dados, un gas contiene siempre el mismo número de partículas, sean átomos o moléculas, independientemente de la naturaleza del gas, haciendo al mismo tiempo la hipótesis de que los gases son moléculas poliatómicas con lo que se comenzó a distinguir entre átomos y moléculas.[16]
El químico ruso Dmítri Ivánovich Mendeléyev creó en 1869 una clasificación de los elementos químicos en orden creciente de su masa atómica, remarcando que existía una periodicidad en las propiedades químicas. Este trabajo fue el precursor de la tabla periódica de los elementos como la conocemos actualmente.[17]
La visión moderna de su estructura interna tuvo que esperar hasta el experimento de Rutherford en 1911 y el modelo atómico de Bohr. Posteriores descubrimientos científicos, como la teoría cuántica, y avances tecnológicos, como el microscopio electrónico, han permitido conocer con mayor detalle las propiedades físicas y químicas de los átomos.[18]

Los elementos básicos de la materia son tres.

Cuadro general de las partículas, quarks y leptones.

Diferencia entre los bariones y los mesones.

Diferencia entre fermiones y bosones.

Tamaño relativo de las diferentes partículas atómicas.
La concepción del átomo que se ha tenido a lo largo de la historia ha
variado de acuerdo a los descubrimientos realizados en el campo de la
física y la química. A continuación se hará una exposición de los
modelos atómicos propuestos por los científicos de diferentes épocas.
Algunos de ellos son completamente obsoletos para explicar los fenómenos
observados actualmente, pero se incluyen a manera de reseña histórica.

Funciones de onda de los primeros orbitales atómicos.

Modelo atómico de Thomson.
Luego del descubrimiento del electrón en 1897 por Joseph John Thomson, se determinó que la materia se
componía de dos partes, una negativa y una positiva. La parte negativa
estaba constituida por electrones, los cuales se encontraban según este
modelo inmersos en una masa de carga positiva a manera de pasas en un
pastel (de la analogía del inglés plum-pudding model) o uvas en
gelatina. Posteriormente Jean
Perrin propuso un modelo modificado a partir del de Thomson donde
las «pasas» (electrones) se situaban en la parte exterior del «pastel»
(la carga positiva).

Modelo atómico de Rutherford.
Este modelo fue desarrollado por el físico Ernest Rutherford a partir de los resultados obtenidos
en lo que hoy se conoce como el experimento de Rutherford en 1911.
Representa un avance sobre el modelo de Thomson, ya que mantiene que el
átomo se compone de una parte positiva y una negativa, sin embargo, a
diferencia del anterior, postula que la parte positiva se concentra en
un núcleo, el cual también contiene virtualmente toda la masa del átomo,
mientras que los electrones se ubican en una corteza orbitando al
núcleo en órbitas circulares o elípticas con un espacio vacío entre
ellos. A pesar de ser un modelo obsoleto, es la percepción más común del
átomo del público no científico.
Rutherford predijo la existencia del neutrón en el año 1920, por esa razón en el modelo anterior (Thomson), no se habla de éste.
Por desgracia, el modelo atómico de Rutherford presentaba varias incongruencias:
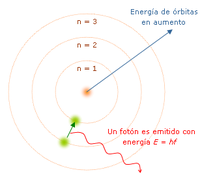
Modelo atómico de Bohr.
Este modelo es estrictamente un modelo del átomo de hidrógeno tomando
como punto de partida el modelo de Rutherford, Niels
Bohr trata de incorporar los fenómenos de absorción y emisión de los
gases, así como la nueva teoría de la cuantización de la energía
desarrollada por Max Planck y el fenómeno del efecto fotoeléctrico observado por Albert Einstein.
«El átomo es un pequeño sistema solar con un núcleo en el centro y electrones moviéndose alrededor del núcleo en órbitas bien definidas». Las órbitas están cuantizadas (los e- pueden estar solo en ciertas órbitas)
Bohr no puede explicar la existencia de órbitas estables y para la condición de cuantización.
Bohr encontró que el momento angular del electrón es h/2π por un método que no puede justificar.

Densidad de probabilidad de ubicación de un electrón para los primeros
niveles de energía.
Después de que Louis-Victor de Broglie propuso la naturaleza
ondulatoria de la materia en 1924, la cual
fue generalizada por Erwin Schrödinger en 1926, se
actualizó nuevamente el modelo del átomo.
En el modelo de Schrödinger se abandona la concepción de los electrones como esferas diminutas con carga que giran en torno al núcleo, que es una extrapolación de la experiencia a nivel macroscópico hacia las diminutas dimensiones del átomo. En vez de esto, Schrödinger describe a los electrones por medio de una función de onda, el cuadrado de la cual representa la probabilidad de presencia en una región delimitada del espacio. Esta zona de probabilidad se conoce como orbital. La gráfica siguiente muestra los orbitales para los primeros niveles de energía disponibles en el átomo de hidrógeno.
‘indivisible’, en realidad está formado por varias partículas subatómicas. El átomo contiene protones, neutrones y electrones, con la excepción del hidrógeno-1, que no contiene neutrones, y del ion hidronio, que no contiene electrones. Los protones y neutrones del átomo se denominan nucleones, por formar parte del núcleo atómico.
El electrón
es la partícula más ligera de cuantas componen el átomo, con una masa de 9,11 · 10−31 kg. Tiene una carga eléctrica negativa cuya magnitud se define como la carga eléctrica elemental, y se ignora si posee subestructura, por lo que se lo considera una partícula elemental. Los protones tienen una masa de 1,67 · 10−27 kg, 1836 veces la del electrón, y una carga positiva opuesta a la de este. Los neutrones tienen un masa de 1,69 · 10−27 kg, 1839 veces la del electrón, y no poseen carga eléctrica. Las masas de ambos nucleones son ligeramente inferiores dentro del núcleo, debido a la energía potencial del mismo; y sus tamaños son similares, con un radio del orden de 8 · 10-16 m o 0,8 femtómetros (fm).[4]
El protón y el neutrón
no son partículas elementales, sino que constituyen un estado ligado de quarks u y d, partículas fundamentales recogidas en el modelo estándar de la física de partículas, con cargas eléctricas iguales a +2/3 y −1/3 respectivamente, respecto de la carga elemental. Un protón contiene dos quarks u y un quark d, mientras que el neutrón contiene dos d y un u, en consonancia con la carga de ambos. Los quarks se mantienen unidos mediante la fuerza nuclear fuerte, mediada por gluones —del mismo modo que la fuerza electromagnética está mediada por fotones—. Además de estas, existen otras partículas subatómicas en el modelo estándar: más tipos de quarks, leptones cargados (similares al electrón), etc.
El núcleo atómico
Los protones y neutrones de un átomose encuentran ligados en el núcleo atómico, la parte central del mismo. El volumen del núcleo es aproximadamente proporcional al número total de nucleones, el número másico A,[5] lo cual es mucho menor que el tamaño del átomo, cuyo radio es del orden de 105 fm o 1 ångström (Å). Los nucleones se mantienen unidos mediante la fuerza nuclear, que es mucho más intensa que la fuerza electromagnética a distancias cortas, lo cual permite vencer la repulsión eléctrica entre los protones.[6]
Los átomos de un
mismo elemento tienen el mismo número de protones, que se denomina
número atómico y se representa por Z. Los átomos de un elemento dado pueden tener distinto número de neutrones: se dice entonces que son isótopos. Ambos números conjuntamente determinan el núclido.
El núcleo atómico puede verse alterado por procesos muy
energéticos en comparación con las reacciones químicas. Los núcleos inestables sufren desintegraciones que pueden cambiar su número de protones y neutrones emitiendo radiación. Un núcleo pesado puede fisionarse en otros más ligeros en una reacción nuclear o espontáneamente. Mediante una cantidad suficiente de energía, dos o más núcleos pueden fusionarse en otro más pesado.
En átomos con
número atómico bajo, los núcleos con una cantidad distinta de protones y neutrones tienden a desintegrarse en núcleos con proporciones más parejas, más estables. Sin embargo, para valores mayores del número atómico, la repulsión mutua de los protones requiere una proporción mayor de neutrones para estabilizar el núcleo.[7]
Nube de electrones

orbitales atómicos.
en el átomo son atraídos por los protones a través de la fuerza electromagnética. Esta fuerza los atrapa en un pozo de potencial electrostático alrededor del núcleo, lo que hace necesaria una fuente de energía externa para liberarlos. Cuanto más cerca está un electrón del núcleo, mayor es la fuerza atractiva, y mayor por tanto la energía necesaria para que escape.
Los electrones, como
otras partículas, presentan simultáneamente propiedades de partícula puntual y de onda, y tienden a formar un cierto tipo de onda estacionaria alrededor del núcleo, en reposo respecto de este. Cada una de estas ondas está caracterizada por un orbital atómico, una función matemática que describe la probabilidad de encontrar al electrón en cada punto del espacio. El conjunto de estos orbitales es discreto, es decir, puede enumerarse, como es propio en todo sistema cuántico. La nube de electrones es la región ocupada por estas ondas, visualizada como una densidad de carga negativa alrededor del núcleo.
Cada orbital corresponde
a un posible valor de energía para los electrones, que se reparten entre ellos. El principio de exclusión de Pauli prohibe que más de dos electrones se encuentren en el mismo orbital. Pueden ocurrir transiciones entre los distintos niveles de energía: si un electrón absorbe un fotón con energía suficiente, puede saltar a un nivel superior; también desde un nivel más alto puede acabar en un nivel inferior, radiando el resto de la energía en un fotón. Las energías dadas por las diferencias entre los valores de estos niveles son las que se observan en las líneas espectrales del átomo.
Propiedades atómicas
Masa
La mayor parte de la masa del átomo viene de los nucleones, los protones y neutrones del núcleo. También contribuyen en una pequeña parte la masa de los electrones, y la energía de ligadura de los nucleones, en virtud de la equivalencia entre masa y energía. La unidad de masa que se utiliza habitualmente para expresarla es la unidad de masa atómica (u). Esta se define como la doceava parte de la masa de un átomo neutro de carbono-12 libre, cuyo núcleo contiene 6 protones y 6 neutrones, y equivale a 1,66 · 10-27 kg aproximadamente. En comparación el protón y el neutrón libres tienen una masa de 1,007 y 1,009 u. La masa de un átomo es entonces aproximadamente igual al número de nucleones en su núcleo —el número másico— multiplicado por la unidad de masa atómica. El átomo estable más pesado es el plomo-208, con una masa de 207,98 u.[8]En química se utiliza también el mol como unidad de masa. Un mol de átomos de cualquier elemento equivale siempre al mismo número de estos (6,022 · 1023), lo cual implica que un mol de átomos de un elemento con masa atómica de 1 u pesa aproximadamente 1 gramo. En general, un mol de átomos de un cierto elemento pesa de forma aproximada tantos gramos como la masa atómica de dicho elemento.
Tamaño
Los átomos no están delimitados por una frontera clara, por lo que su tamaño se equipara con el de su nube electrónica. Sin embargo, tampoco puede establecerse una medida de esta, debido a las propiedades ondulatorias de los electrones. En la práctica, se define el radio atómico estimándolo en función de algún fenómeno físico, como la cantidad y densidad de átomos en un volumen dado, o la distancia entre dos núcleos en una molécula.Los diversos métodos existentes arrojan valores para el radio atómico de entre 0,5 y 5 Å. Dentro de la tabla periódica de los elementos, el tamaño de los átomos tiende a disminuir a lo largo de un periodo —una fila—, para aumentar súbitamente al comienzo de uno nuevo, a medida que los electrones ocupan niveles de energía más altos.[9]
Las dimensiones del átomo son miles de veces más pequeñas que la longitud de onda de la luz (400-700 nm) por lo que estos no pueden ser observados utilizando instrumentos ópticos. En comparación, el grosor de un cabello humano es equivalente a un millón de átomos de carbono. Si una manzana fuera del tamaño de la Tierra, los átomos en ella serían tan grandes como la manzana original.[10]
Niveles de energía
Un electrón ligado en el átomo posee una energía potencial inversamente proporcional a su distancia al núcleo y de signo negativo, lo que quiere decir que esta aumenta con la distancia. La magnitud de esta energía es la cantidad necesaria para desligarlo, y la unidad usada habitualmente para expresarla es el electrónvoltio (eV). En el modelo mecanocuántico solo hay un conjunto discreto de estados o niveles en los que un electrón ligado puede encontrarse —es decir, enumerables—, cada uno con un cierto valor de la energía. El nivel con el valor más bajo se denomina el estado fundamental, mientras que el resto se denominan estados excitados.Cuando un electrón efectúa una transición entre dos estados distintos, absorbe o emite un fotón, cuya energía es precisamente la diferencia entre los dos niveles. La energía de un fotón es proporcional a su frecuencia, así que cada transición se corresponde con una banda estrecha del espectro electromagnético denominada línea espectral.

Algunas líneas espectrales se presentan muy juntas entre sí, tanto que llegaron a confundirse con una sola históricamente, hasta que fue descubierta su subestructura o estructura fina. La causa de este fenómeno se encuentra en las diversas correcciones a considerar en la interacción entre los electrones y el núcleo. Teniendo en cuenta tan solo la fuerza electrostática, ocurre que algunas de las configuraciones electrónicas pueden tener la misma energía aun siendo distintas. El resto de pequeños efectos y fuerzas en el sistema electrón-núcleo rompe esta redundancia o degeneración, dando lugar a la estructura fina. Estos incluyen las correcciones relativistas al movimiento de electrón, la interacción de su momento magnético con el campo eléctrico y con el núcleo, etc.[11]
Además, en presencia de un campo externo los niveles de energía se ven modificados por la interacción del electrón con este, en general produciendo o aumentando la división entre los niveles de energía. Este fenómeno se conoce como efecto Stark en el caso de un campo eléctrico, y efecto Zeeman en el caso de un campo magnético.
Las transiciones de un electrón a un nivel superior ocurren en presencia de radiación electromagnética externa, que provoca la absorción del fotón necesario. Si la frecuencia de dicha radiación es muy alta, el fotón es muy energético y el electrón puede liberarse, en el llamado efecto fotoeléctrico.
Las transiciones a un nivel inferior pueden ocurrir de manera espontánea, emitiendo la energía mediante un fotón saliente; o de manera estimulada, de nuevo en presencia de radiación. En este caso, un fotón «entrante» apropiado provoca que el electrón decaiga a un nivel con una diferencia de energía igual a la del fotón entrante. De este modo, se emite un fotón saliente cuya onda asociada está sincronizada con la del primero, y en la misma dirección. Este fenómeno es la base del láser.
Interacciones eléctricas entre protones y electrones
Antes del experimento de Rutherford la comunidad científica aceptaba el modelo atómico de Thomson, situación que varió después de la experiencia de Rutherford. Los modelos posteriores se basan en una estructura de los átomos con una masa central cargada positivamente rodeada de una nube de carga negativa.[12]Este tipo de estructura del átomo llevó a Rutherford a proponer su modelo en que los electrones se moverían alrededor del núcleo en órbitas. Este modelo tiene una dificultad proveniente del hecho de que una partícula cargada acelerada, como sería necesario para mantenerse en órbita, radiaría radiación electromagnética, perdiendo energía. Las leyes de Newton, junto con las ecuaciones de Maxwell del electromagnetismo aplicadas al átomo de Rutherford llevan a que en un tiempo del orden de 10−10 s, toda la energía del átomo se habría radiado, con la consiguiente caída de los electrones sobre el núcleo.[13]
Historia de la teoría atómica
El concepto de átomo existe desde la Antigua Grecia propuesto por los filósofos griegos Demócrito, Leucipo y Epicuro, sin embargo, no se generó el concepto por medio de la experimentación sino como una necesidad filosófica que explicara la realidad, ya que, como proponían estos pensadores, la materia no podía dividirse indefinidamente, por lo que debía existir una unidad o bloque indivisible e indestructible que al combinarse de diferentes formas creara todos los cuerpos macroscópicos que nos rodean.[14] El siguiente avance significativo no se realizó hasta que en 1773 el químico francés Antoine-Laurent de Lavoisier postuló su enunciado: «La materia no se crea ni se destruye, simplemente se transforma». La ley de conservación de la masa o ley de conservación de la materia; demostrado más tarde por los experimentos del químico inglés John Dalton quien en 1804, luego de medir la masa de los reactivos y productos de una reacción, y concluyó que las sustancias están compuestas de átomos esféricos idénticos para cada elemento, pero diferentes de un elemento a otro.[15]Luego en 1811, el físico italiano Amedeo Avogadro, postuló que a una temperatura, presión y volumen dados, un gas contiene siempre el mismo número de partículas, sean átomos o moléculas, independientemente de la naturaleza del gas, haciendo al mismo tiempo la hipótesis de que los gases son moléculas poliatómicas con lo que se comenzó a distinguir entre átomos y moléculas.[16]
El químico ruso Dmítri Ivánovich Mendeléyev creó en 1869 una clasificación de los elementos químicos en orden creciente de su masa atómica, remarcando que existía una periodicidad en las propiedades químicas. Este trabajo fue el precursor de la tabla periódica de los elementos como la conocemos actualmente.[17]
La visión moderna de su estructura interna tuvo que esperar hasta el experimento de Rutherford en 1911 y el modelo atómico de Bohr. Posteriores descubrimientos científicos, como la teoría cuántica, y avances tecnológicos, como el microscopio electrónico, han permitido conocer con mayor detalle las propiedades físicas y químicas de los átomos.[18]
Evolución del modelo atómico





Modelo de Dalton
Fue el primer modelo atómico con bases científicas, fue formulado en 1808 por John Dalton, quien imaginaba a los átomos como diminutas esferas.[19] Este primer modelo atómico postulaba:- La materia está formada por partículas muy pequeñas llamadas átomos, que son indivisibles y no se pueden destruir.
- Los átomos de un mismo elemento son iguales entre sí, tienen su propio peso y cualidades propias. Los átomos de los diferentes elementos tienen pesos diferentes.
- Los átomos permanecen sin división, aun cuando se combinen en las reacciones químicas.
- Los átomos, al combinarse para formar compuestos guardan relaciones simples.
- Los átomos de elementos diferentes se pueden combinar en proporciones distintas y formar más de un compuesto.
- Los compuestos químicos se forman al unirse átomos de dos o más elementos distintos.
Modelo de Thomson


Detalles del modelo atómico
Para explicar la formación de iones, positivos y negativos, y la presencia de los electrones dentro de la estructura atómica, Thomson ideó un átomo parecido a un pastel de frutas. Una nube positiva que contenía las pequeñas partículas negativas (los electrones) suspendidos en ella. El número de cargas negativas era el adecuado para neutralizar la carga positiva. En el caso de que el átomo perdiera un electrón, la estructura quedaría positiva; y si ganaba, la carga final sería negativa. De esta forma, explicaba la formación de iones; pero dejó sin explicación la existencia de las otras radiaciones.Modelo de Rutherford

Rutherford predijo la existencia del neutrón en el año 1920, por esa razón en el modelo anterior (Thomson), no se habla de éste.
Por desgracia, el modelo atómico de Rutherford presentaba varias incongruencias:
- Contradecía las leyes del electromagnetismo de James Clerk Maxwell, las cuales estaban muy comprobadas mediante datos experimentales. Según las leyes de Maxwell, una carga eléctrica en movimiento (en este caso el electrón) debería emitir energía constantemente en forma de radiación y llegaría un momento en que el electrón caería sobre el núcleo y la materia se destruiría. Todo ocurriría muy brevemente.
- No explicaba los espectros atómicos.
Modelo de Bohr

«El átomo es un pequeño sistema solar con un núcleo en el centro y electrones moviéndose alrededor del núcleo en órbitas bien definidas». Las órbitas están cuantizadas (los e- pueden estar solo en ciertas órbitas)
- Cada órbita tiene una energía asociada. La más externa es la de mayor energía.
- Los electrones no radian energía (luz) mientras permanezcan en órbitas estables.
- Los electrones pueden saltar de una a otra órbita. Si lo hace desde una de menor energía a una de mayor energía absorbe un cuanto de energía (una cantidad) igual a la diferencia de energía asociada a cada órbita. Si pasa de una de mayor a una de menor, pierde energía en forma de radiación (luz).
Bohr no puede explicar la existencia de órbitas estables y para la condición de cuantización.
Bohr encontró que el momento angular del electrón es h/2π por un método que no puede justificar.
Modelo de Schrödinger

En el modelo de Schrödinger se abandona la concepción de los electrones como esferas diminutas con carga que giran en torno al núcleo, que es una extrapolación de la experiencia a nivel macroscópico hacia las diminutas dimensiones del átomo. En vez de esto, Schrödinger describe a los electrones por medio de una función de onda, el cuadrado de la cual representa la probabilidad de presencia en una región delimitada del espacio. Esta zona de probabilidad se conoce como orbital. La gráfica siguiente muestra los orbitales para los primeros niveles de energía disponibles en el átomo de hidrógeno.
Molécula
Casi toda la química orgánicay buena
parte de la química inorgánica se ocupan de la síntesis y reactividad de moléculas y compuestos moleculares. La química física y, especialmente, la química cuántica también estudian, cuantitativamente, en su caso, las propiedades y reactividad de las moléculas. La bioquímica está íntimamente relacionada con la biología molecular, ya que ambas estudian a los seres vivos a nivel molecular. El estudio de las interacciones específicas entre moléculas, incluyendo el reconocimiento molecular es el campo de estudio de la química supramolecular. Estas fuerzas explican las propiedades físicas como la solubilidad o el punto de ebullición de un compuesto molecular.
Las moléculas rara vez se
encuentran sin interacción entre ellas, salvo en gases enrarecidos. Así, pueden encontrarse en redes cristalinas, como el caso de las moléculas de H2O en el hielo o con interacciones intensas pero que cambian rápidamente de direccionalidad, como en el agua líquida. En orden creciente de intensidad, las fuerzas intermoleculares más relevantes son: las fuerzas de Van der Waals y los puentes de hidrógeno. La dinámica molecular es un método de simulación por computadora que utiliza estas fuerzas para tratar de explicar las propiedades de las moléculas.
Tipos de moléculas
Las moléculas se pueden clasificar en:- Moléculas discretas, constituidas
- por un número bien definido de átomos, sean estos del mismo elemento (moléculas homonucleares, como el dinitrógeno o el fullereno) o de elementos distintos (moléculas heteronucleares, como el agua).
-
 Molécula de agua, "disolvente
Molécula de agua, "disolvente
universal", de importancia fundamental en innumerables procesos bioquímicos e industriales -
 Representación poliédrica del anión de Keggin, un polianión molecular
Representación poliédrica del anión de Keggin, un polianión molecular




- Macromoléculas o polímeros, constituidas por la repetición
- de una unidad comparativamente simple -o un conjunto limitado de dichas unidades- y que alcanzan pesos moleculares relativamente altos.
-

-
 Representación de un fragmento lineal de polietileno, el plástico más usado
Representación de un fragmento lineal de polietileno, el plástico más usado -
 Primera generación de un dendrímero, un tipo especial de polímero que crece de forma fractal
Primera generación de un dendrímero, un tipo especial de polímero que crece de forma fractal


Descripción
La estructura molecularpuede ser descrita de diferentes formas. La fórmula química es útil para moléculas sencillas, como H2O para el agua o NH3 para el amoníaco. Contiene los símbolos de los elementos presentes en la molécula, así como su proporción indicada por los subíndices.
Para moléculas más complejas,
como las que se encuentran comúnmente en química orgánica, la fórmula química no es suficiente, y vale la pena usar una fórmula estructural o una fórmula esqueletal, las que indican gráficamente la disposición espacial de los distintos grupos funcionales.
Cuando se quieren mostrar variadas propiedades moleculares...
(como el potencial eléctrico en la superficie de la molécula), o se trata de sistemas muy complejos, como proteínas, ADN o polímeros, se utilizan representaciones especiales, como los modelos tridimensionales (físicos o representados por ordenador). En proteínas, por ejemplo, cabe distinguir entre estructura primaria (orden de los aminoácidos), secundaria (primer plegamiento en hélices, hojas, giros...), terciaria (plegamiento de las estructuras tipo hélice/hoja/giro para dar glóbulos) y cuaternaria (organización espacial entre los diferentes glóbulos).

Figura 1. Representaciones
de la terpenoide, atisano, 3D
(centro izquierda) y 2D (derecha). En el modelo 3D de la
izquierda, los átomos de carbono están representados por esferas grises; las
blancas representan a los átomos de hidrógeno
y los cilindros representan los enlaces. El modelo es una
representación de la superficies molecular, coloreada por áreas de carga eléctrica positiva (rojo) o negativa (azul). En el
modelo 3D del centro, las esferas azul claro representan átomos de
carbono, las blancas de hidrógeno y los cilindros entre los átomos son
los enlaces simples.
Observación
de epidermis de cebolla
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
MATERIAL
TÉCNICA
El resultado de esta práctica debe ser semejante a lo que muestran las siguientes fotos obtenidas con microscopio óptico:
MoneraMonera[1] es un reino de la clasificación de los seres vivos, considerado actualmente obsoleto por la mayoría de especialistas. En la influyente clasificación de Margulis, significa lo mismo que procariotas, y así sigue siendo usada en muchos manuales y libros de texto.Este reino comprende entre 4.000 y 9.000 especies que habitan todos los ambientes. Son organismos microscópicos, formados por una sola célula sin núcleo. Abarca dos grupos importantes: arqueobacterias y eubacterias (que incluye las cianobacterias). 
aceptada internacionalmente. Este Reino lo integran no sólo las bacterias, también pertenecen a él las llamadas algas verde azuladas. Este Reino, rico en diversidad de especies, agrupa a los organismos denominados procariontes por poseer un tipo de célula nombrada procariota carentes de núcleo rodeado por membrana. En estudios de laboratorio se determinó que tienen sólo un cromosoma circular y ribosomas que sedimentan a 70S mientras que los eucariotas lo hacen a 80S. Al ser observadas por el microscopio electrónico se corrobora la ausencia de organelas rodeadas por membranas. A los miembros de este reino se les consideran los primeros pobladores o formas de vida sobre la Tierra, investigaciones científicas sostienen que existían desde hace unos 3.500.000.000 años es decir que pudieron ser reinantes en la atmósfera primitiva. La formación del oxígeno limitó a las bacterias a ambientes anaerobios (metanógenas) o con condiciones extremas (bacterias halófilas y bacterias termoacidófilas). Los Moneras son unicelulares, autótrofos o heterótrofos, se pueden encontrar en todos los ambientes, algunas bacterias se encuentran en lugares realmente sorprendentes como las bacterias termófilas, las bacterias pueden ser inmóviles o móviles, presentan la pared rígida y su reproducción es asexual, aunque pueden tener intercambios de información genética entre los individuos de una misma especie o de especies distintas. Donde todavía se usa, el término Monera designa un grado (nivel evolutivo) formado por los organismos celulares que carecen de núcleo bien definido, los que son llamados procariontes y que son considerados las formas de vida más antiguas. Características generales:
ClasificaciónTradicionalmente el reino Monera se clasificaba durante el siglo XX hasta los años 1970 en dos grandes grupos o divisiones: bacterias y cianobacterias (cianofíceas o algas azul-verdosas). A su vez las bacterias se subclasificaban según su morfología, tal como lo hacían las clasificaciones del siglo XIX. Un avance importante en clasificación procariota significaron las del Manuel de Bergay de 1978 y 1984 atribuidas sobre todo a R.G.E. Murray, las cuales se basaron principalmente en la estructura de pared y membranas celulares, procurando además evitar nombres en latín en donde se sabía a conciencia que era imposible determinar las verdaderas relaciones filogenéticas; o la clasificación de Margulis y Schwartz de 1982 basada en metabolismo y bioquímica bacteriana.Pero la verdadera revolución vino del descubrimiento del análisis del ARN ribosomal 16S y 5S desarrollado por C. Woese, el cual fue el más grande avance en taxonomía procariota desde el descubrimiento de la tinción de Gram en 1884 y permitió al fin integrar en forma real el análisis filogenético a la microbiología, el cual era aplicable casi exclusivamente a plantas y animales. A continuación se muestra una síntesis del reino Monera o Procaryotae de inicios de los 1980:. FungiEn biología, el término Fungi (latín, literalmente "hongos") designa a un grupo de organismos eucariotas entre los que se encuentran los mohos, las levaduras y las setas. Se clasifican en un reino distinto al de las plantas, animales y bacterias. Esta diferenciación se debe, entre otras cosas, a que poseen paredes celulares compuestas por quitina, a diferencia de las plantas, que contienen celulosa y debido a que algunos crecen y/o actúan como parásitos de otras especies. Actualmente se consideran como un grupo heterogéneo, polifilético, formado por organismos pertenecientes por lo menos a tres líneas evolutivas independientes. Los hongos se encuentran en hábitats muy diversos: pueden ser pirófilos (Pholiota carbonaria) o coprófilos (Psilocybe coprophila). Según su ecología, se pueden clasificar en cuatro grupos: saprofitos, liquenizados, micorrizógenos y parásitos. Los hongos saprofitos pueden ser sustrato específicos: Marasmius buxi o no específicos: Mycena pura. Los simbiontes pueden ser: hongos liquenizados Basidiolichenes: Omphalina ericetorum y ascolichenes: Cladonia coccifera y hongos micorrízicos: específicos: Lactarius torminosus (solo micorriza con abedules) y no específicos: Hebeloma mesophaeum. En la mayoría de los casos, sus representantes son poco conspicuos debido a su diminuto tamaño; suelen vivir en suelos y juntos a materiales en descomposición y como simbiontes de plantas, animales u otros hongos. Cuando fructifican, no obstante, producen esporocarpos llamativos (las setas son un ejemplo de ello). Realizan una digestión externa de sus alimentos, secretando enzimas, y que absorben luego las moléculas disueltas resultantes de la digestión. A esta forma de alimentación se le llama osmotrofia, la cual es similar a la que se da en las plantas, pero, a diferencia de aquéllas, los nutrientes que toman son orgánicos. Los hongos son los descomponedores primarios de la materia muerta de plantas y de animales en muchos ecosistemas, y como tales poseen un papel ecológico muy relevante en los ciclos biogeoquímicos. Los hongos tienen una gran importancia económica: las levaduras son las responsables de la fermentación de la cerveza y el pan, y se da la recolección y el cultivo de setas como las trufas. Desde 1940 se han empleado para producir industrialmente antibióticos, así como enzimas (especialmente proteasas). Algunas especies son agentes de biocontrol de plagas. Otras producen micotoxinas, compuestos bioactivos (como los alcaloides) que son tóxicos para humanos y otros animales. Las enfermedades fúngicas afectan a humanos, otros animales y plantas; en estas últimas, afecta a la seguridad alimentaria y al rendimiento de los cultivos. Los hongos se presentan bajo dos formas principales: hongos filamentosos (antiguamente llamados "mohos") y hongos levaduriformes. El cuerpo de un hongo filamentoso tiene dos porciones, una reproductiva y otra vegetativa.[1] La parte vegetativa, que es haploide y generalmente no presenta coloración, está compuesta por filamentos llamados hifas (usualmente microscópicas); un conjunto de hifas conforma el micelio[2] (usualmente visible). A menudo las hifas están divididas por tabiques llamados septos. Los hongos levaduriformes —o simplemente levaduras— son siempre unicelulares, de forma casi esférica. No existen en ellos una distinción entre cuerpo vegetativo y reproductivo. Dentro del esquema de los cinco reinos de Wittaker y Margulis, los hongos pertenecen en parte al reino protista (los hongos ameboides y los hongos con zoosporas) y al reino Fungi (el resto). En el esquema de ocho reinos de Cavalier-Smith pertenecen en parte al reino Protozoa (los hongos ameboides), al reino Chromista (los Pseudofungi) y al reino Fungi todos los demás.. La diversidad de taxa englobada en el grupo está poco estudiada; se estima que existen unas 1,5 millones de especies, de las cuales apenas el 5% han sido clasificadas. Durante los siglos XVIII y XIV, Linneo, Christian Hendrik Persoon, y Elias Magnus Fries clasificaron a los hongos de acuerdo a su morfología o fisiología. Actualmente, las técnicas de Biología Molecular han permitido el establecimiento de una taxonomía molecular basada en secuencias de ácido desoxirribonucleico (ADN), que divide al grupo en siete filos. La especialidad de la medicina y de la botánica que se ocupa de los hongos se llama micología, donde se emplea el sufijo -mycota para las divisiones y -mycetes para las clases.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||


























{kind=link}
No hay comentarios:
Publicar un comentario